Morfología de la lámina aracnoidea espinal humana.
M. A. Reina , A. Prats-Galino A , R. G. Sola , A. Puigdellívol-Sánchez , R. Arriazu Navarro , J. A. De Andrés:
1 Departamento de Anestesiología. Hospital Universitario Madrid Montepríncipe. Departamento de Ciencias Médicas Clínicas e Instituto de Medicina Molecular Aplicada. Facultad de Medicina, Universidad CEU San Pablo. Madrid.
2 Unidad de Anatomía y Embriología Humana. Facultad de Medicina. Universidad de Barcelona, Barcelona.
3 Servicio de Neurocirugía. Hospital Universitario La Princesa. Universidad Autónoma de Madrid. Unidad de Neurocirugía. Hospital Universitario Madrid Montepríncipe.
4 Unidad de Histología Humana. Departamento de Ciencias Médicas Básicas e Instituto de Medicina Molecular Aplicada. Facultad de Medicina, Universidad CEU San Pablo. Madrid.
5 Departamento de Anestesiología. Hospital General Universitario. Valencia.
Morfología de la lámina aracnoidea espinal humana. Barrera que limita la permeabilidad del saco dural.
Resumen
Objetivos : Se ha demostrado que las moléculas inyectadas en el espacio epidural pasan desde éste al espacio subaracnoideo por difusión simple a través de la pared del saco dural. Nuestro objetivo fue estudiar la ultraestructura de células de la lámina aracnoidea y tipo de uniones especializadas responsables del efecto barrera que gobierna el tránsito de moléculas a través del saco dural humano.
Material y método : Se estudiaron catorce muestras de la lámina aracnoidea obtenidas de dos pacientes durante intervenciones con apertura del saco dural lumbar. Las muestras se trataron con glutaraldehido, tetróxido de osmio, ferrocianuro, acetona, e incluyeron en resina. Los cortes ultrafinos se contrastaron con citrato de plomo, para poder ser observados con un microscopio electrónico de transmisión.
Resultados : La lámina aracnoidea posee un espesor de 35-40 μm. En su porción externa se hallan células neuroteliales del compartimento subdural, mientras que su porción interna está formada por un plano celular de 5-8 μm de espesor, constituido por la superposición de 4-5 células aracnoideas que forman la capa barrera. El espacio intercelular de este plano fue de 0,02-0,03 μm. Entre las células aracnoideas se encontraron uniones especializadas de membrana de tipo desmosomas y uniones estrechas.
Conclusiones : Las células aracnoideas poseen características estructurales que aseguran la función barrera del saco dural humano y no ocupan todo el espesor de la lámina aracnoidea, sólo su porción interna. La presencia de uniones especializadas de membrana entre sus células justifica la permeabilidad selectiva de esta lámina.
Palabras clave:
Duramadre. Aracnoides. Permeabilidad. Anestesia epidural. Ultraestructura. Histología.
Correspondencia:
Dr. M. A. Reina Dpto. de Anestesiología Hospital Universitario Montepríncipe Avda. Montepríncipe, 25 28660 Boadilla del Monte (Madrid)
E-mail: miguelangel@perticone.e.telefonica.net Aceptado para su publicación en agosto de 2010.
Structure of the arachnoid layer of the human spinal meninges: a barrier that regulates dural sac permeability
Summary
Objetives : Drugs injected into the epidural space are known to penetrate the subarachnoid space by simple diffusion through the dural sac. We aimed to study the cellular ultrastructure of the arachnoid membrane and the type of intercellular junctions responsible for creating the barrier that regulates the passage of drugs through the dural sac in humans.
Material and methods : Fourteen tissue samples of arachnoid membrane were taken from 2 patients during procedures that required opening the lumbar dural sac. The samples were treated with glutaraldehyde, osmium tetroxide, ferrocyanide and acetone, and then embedded in resin. Ultrathin sections were stained with lead citrate for examination by transmission electron microscopy.
Results : The arachnoid membrane was 35 to 40 μm thick. The outer surface contained neurothelial cells (dural border cells) along the subdural compartment, while the internal portion was made up of a plane 5 to 8 μm thick with 4 to 5 arachnoid cells overlapping to form a barrier layer. The intercellular spaces on this plane were 0.02 to 0.03 μm wide; the arachnoid cells were bridged by specialized junctions (desmosomes and other tight junctions).
Conclusions : Structural features of the arachnoid cells provide a barrier within the human dural sac. They occupy only the internal portion of the arachnoid membrane. Specialized intercellular junctions explain the selective permeability of this membrane.
Key words:
Dura mater. Arachnoid membrane: permeability. Epidural anesthesia.
Introducción
Durante años se ha mantenido que, tras la administración de un fármaco en el espacio epidural, se produce un paso hacia el espacio subaracnoideo, en parte, a través de las vellosidades aracnoideas localizadas en los manguitos durales 1-4 y, en parte, por difusión a través del espesor del saco dural.
La hipótesis del paso de moléculas a través de las vellosidades aracnoideas se basaba probablemente en la idea inversa: el tránsito de sustancias desde el líquido cefalorraquídeo (LCR) hacia el espacio epidural. Este tránsito había sido propuesto por Key y Retzius 5 a finales del siglo XIX, y más tarde, confirmado de forma experimental en animales 6,7 . Sin embargo, este mecanismo es unidireccional, no bidireccional 8 . Las vellosidades aracnoideas están formadas por células que protruyen en el espesor de la duramadre a diferentes profundidades, algunas de las cuales alcanzan la luz de las venas epidurales. De esta forma, las moléculas contenidas en el espacio subaracnoideo podrían ser transportadas, a través de las células aracnoideas, a la sangre venosa, por mecanismos de pinocitosis del LCR, y posterior exocitosis hacia la sangre venosa 6 . En este caso, el material sería eliminado por los vasos del espacio epidural. Apoyados en esta idea, se propuso el mecanismo inverso, es decir, que los anestésicos locales depositados en el espacio epidural podrían pasar al LCR a través de las vellosidades aracnoideas de los manguitos durales. Las investigaciones realizadas en las últimas dos décadas por Bernards 8,9 parecen descartar esta hipótesis, sugiriendo además que sustancias inyectadas en el espacio epidural no pueden llegar a la médula espinal por difusión a través de las arterias radiculares, sino que sólo pasan al LCR por difusión simple a través del espesor del saco dural 10-12 .
Por esta razón, resulta de especial interés conocer qué estructura del saco dural es la responsable de controlar el paso de moléculas. En estudios previos 13-17 hemos descrito que la duramadre humana está formada por láminas durales concéntricas, cada una de ellas constituida a su vez por fibras de colágeno dispuestas en diferentes direcciones, escasas fibras elásticas con un diámetro 20 veces mayor y algunos fibroblastos. Entre las fibras se halla una sustancia amorfa formada por mucopolisacáridos y proteoglicanos, que permiten un libre paso de sustancias hidrosolubles y convierten a la duramadre en una membrana permeable. De esta forma, la lámina aracnoidea, que ocupa apenas el 10% del espesor interno del saco dural, podría ser la responsable del efecto barrera de sustancias, que desde el espacio epidural pasan al LCR, aunque todavía no sabemos si todo o parte del espesor de la lámina aracnoidea es la responsable.
Vandenabeele et al 18 , en 1996, estudiaron la duramadre y la aracnoides espinal por microscopía electrónica de transmisión en muestras extraídas durante intervenciones quirúrgicas de la columna vertebral y, en estudios previos, nosotros describimos, en cadáveres recién fallecidos, la duramadre 13-17 , las células neuroteliales del compartimento subdural 19,20 , la aracnoides trabecular 21-23 , la piamadre 24,25 y la ultraestructura de los manguitos durales 26,27 , usando microscopía electrónica de transmisión y de barrido. El objetivo de este trabajo ha sido estudiar la ultraestructura de la lámina aracnoidea, el tipo de uniones especializadas de membrana entre las células aracnoideas, y conocer si, en base a las características morfológicas, todo o parte de esa lámina aracnoidea podría ser la responsable del efecto barrera, en el saco dural humano.
Material y método
Para la realización de este trabajo se ha contado con la aprobación del Comité de Ética de Investigaciones Clínicas (CEIC) del hospital, así como con el consentimiento informado de los pacientes intervenidos quirúrgicamente, para la obtención de muestras de tejidos. Se extrajeron muestras de membrana aracnoidea en dos pacientes, aprovechando el momento de la apertura de la aracnoides y la necesidad de extirparla parcialmente, para permitir una mejor visión y manipulación quirúrgica de la lesión intradural (neurinomas de la cola de caballo).
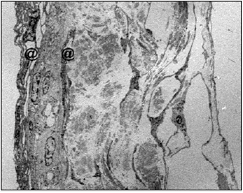
La lámina aracnoidea es una membrana fina y traslúcida que se separa de la duramadre con la manipulación del saco dural. Durante la cirugía, la apertura de la duramadre se realizó manteniendo intacta la lámina aracnoidea, que retiene el LCR en el interior del espacio subaracnoideo, que permite ver por transparencia el contenido del saco dural (Fig. 1). En la r egión lumbar, donde se realizaron las intervenciones quirúrgicas, pudieron observarse por transparencia las raíces nerviosas de la cola de caballo.
La lámina aracnoidea se rompe con mucha facilidad, si no se extreman los cuidados durante las maniobras de apertura de la duramadre. Al abrir esta membrana, es preciso extirparla parcialmente, para facilitar la identificación de la lesión, así como la disección de las raíces y extirpación tumoral mediante técnicas microquirúrgicas. Esos fragmentos, separados del saco dural, fueron las muestras utilizadas para el estudio microscópico.
En total se estudiaron muestras obtenidas de catorce fragmentos, ocho del primer paciente y seis del segundo. Las muestras se fijaron 4 horas en una solución de glutaraldehido al 2,5% en tampón fosfato


(pH = 7,2-7,3) y fueron postfijadas en una solución de tetróxido de osmio al 1% y ferrocianuro al 1% durante 1 hora. Posteriormente se deshidrataron por inmersión en soluciones de acetona de concentraciones crecientes, iniciándose con soluciones de acetona al 30% hasta solución de acetona al 100%. Las muestras se incluyeron en resina epoxi Epon 812, que se polimerizó a 60ºC durante 24 horas. Se obtuvieron cortes semifinos de control con espesores de 0,5 μm, que fueron teñidos con azul de metileno de Richarson. Los cortes ultrafinos, de 70 nanómetros de espesor, obtenidos con un ultramicrotomo Reichert Jung ultracut E (Viena, Austria), se contrastaron con citrato de plomo de Reynold durante 3 minutos. Finalmente, las muestras se observaron con un microscopio electrónico de transmisión TEM Jeol 1010 (JEOL Corp. Ltd., Tokio, Japón). La tinción usada permitió un nivel de contraste suficiente entre estructuras vecinas, necesario para evaluar la ultraestructura celular, la morfología de las uniones especializadas de membrana, y de los espacios intercelulares.
Resultados
Los resultados obtenidos fueron similares en todas las muestras. Del total de muestras se estudiaron once muestras y tres fueron descartadas por artefactos derivados de su procesamiento.
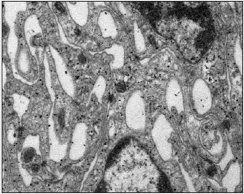
Bajo el microscopio electrónico, esta lámina tenía un espesor de 35-40 μm y estaba constituida por cuatro porciones bien diferenciadas: desde externo hacia interno encontramos una primera porción ocupada por células alargadas, ramificadas y oscuras, próxima al espacio subdural adquirido, que se corresponden con las células neuroteliales del compartimento subdural, también denominadas por otros autores, 18 células del borde dural. Una segunda porción ocupada por fibras de colágeno con diferentes direcciones que podía ocupar entre el 40 y 50% del espesor total de la lámina aracnoidea. A continuación y separado por una membrana basal, ocasionalmente interrumpida, se encontraba una tercera porción ocupada por las células aracnoideas que formaban la capa barrera. Esta capa barrera posee 5-8 μm de espesor y estaba constituida por 4-5 planos de células aracnoidea s (Figs. 2, 3 y 4). Estas células, de coloración clara, tenían morfología alargada con un espesor que varía entre 1,5 a 2 μm. En el citoplasma de estas células se observó una matriz reticular con finos filamentos orientados en diferentes direcciones, retículo endoplásmico, numerosas vesículas y algunos lisosomas y mitocondrias. La longitud de estas células excedía los 40 μm. El núcleo de estas células es de gran tamaño (1 μm de anchura y 7-9 μm de longitud), electrolúcido, y con pequeños grumos de heterocromatina. El espacio intercelular a nivel de la
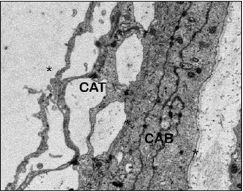
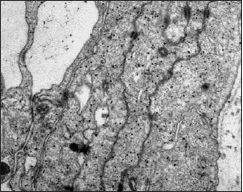
capa barrera era mínimo, formando estrechas hendiduras de 0,02-0,03 μm. Entre las membranas plasmáticas de células vecinas se podían encontrar diferentes tipos de uniones especializadas (Fig. 5), observándose abundantes uniones estrechas (tight junctions) y desmosomas. En este espacio intercelular no había colágeno, fibras elásticas, ni microfibrillas.
Por último, se encontraba la cuarta porción, más interna y en contacto con el LCR ocupada por la aracnoides reticular, donde las células aracnoideas ocupaban una posición interna a la capa barrera y estaban en contacto con el espacio subaracnoideo. Estas células
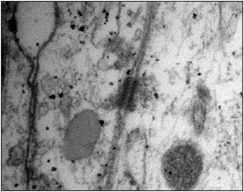
presentaban un progresivo aumento del espacio intercelular (Fig. 6) , facilitado por la falta de uniones especializadas de membrana, dando origen a pequeñas lagunas intercelulares, que aumentaban de tamaño a medida que se progresa hacia el espacio subaracnoideo (Fig. 6).
Discusión
La lámina aracnoidea, entre sus componentes, tiene una capa de células fuertemente unidas entre sí, que puede justificar su efecto barrera, y que hemos denominado capa barrera de la aracnoides. Esta capa (tercera porción) representa por sí sola, aproximadamente el 20% del espesor de la lámina aracnoidea. En la primera porción más externa, se han encontrado células neuroteliales que se rompen durante las maniobras de apertura dural, para dar origen al espacio subdural adquirido. Internamente (segunda porción), se encuentran fibras de colágeno que aportarían resistencia mecánica a la lámina aracnoidea. Más interno, las células aracnoideas de la capa barrera que controlan la difusión, y en la porción más interna y próximo al LCR, las células alargadas con menos uniones especializadas de membrana, que daban origen al componente celular de la aracnoides reticular. En nuestras muestras, esta estructura reticular estaba formada exclusivamente por células, sin presencia de fibras de colágeno. A medida que esa estructura se proyectaba hacia el interior del espacio subaracnoideo, las células aracnoideas rodeaban paquetes de colágeno, y darían origen al trabeculado aracnoideo, estructura con mayor resistencia mecánica, que se extiende hasta vasos y
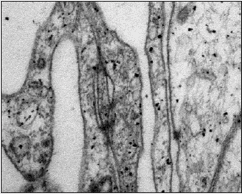
Las uniones especializadas de membrana mantienen fuertemente unidas a las células aracnoideas entre sí, en especial a las células que forman la capa barrera. Por sus características morfológicas, nosotros identificamos uniones tipo desmosomas y tipo estrechas a lo largo de una misma membrana celular. Este tipo de uniones han sido más estudiadas en los epitelios 28-30 . El efecto barrera y la diferente permeabilidad selectiva que ofrecen algunas membranas de naturaleza celular, se debe en parte, a la presencia de moléculas que aportan adherencia celular, y en parte, a las uniones especializadas de membrana. Algunas de las proteínas que contribuyen a la adherencia entre células, son componentes de las mismas uniones intercelulares.
Las uniones estrechas, también llamadas herméticas, tight junctions o zonula occludens, son estructuras complejas que impiden el paso libre de líquidos y solutos entre las células 28-30 . Los componentes moleculares en este tipo de uniones son proteínas de alto peso molecular, organizadas en filas paralelas que permiten mantener unidas dos membranas plasmáticas de células adyacentes. Entre estas proteínas se han identificado la ocludina y claudina.
El grado de permeabilidad a través de estas membranas, estaría relacionado con el número de filas paralelas que ocupen estas proteínas 28-30 . En las zonas de la membrana celular que existe este tipo de unión, las moléculas e iones podrían atravesar las células por vía transcelular, o difundir a través de las uniones, por vía paracelular. El agua difundiría a través de las célu-
las, pero la mayoría de los iones e incluso las moléculas pequeñas, como los azúcares simples y aminoácidos no podrían pasar. El microscopio electrónico permite observar que en las zonas de estas uniones, las membranas celulares se tocan, o incluso se funden, sin dejar espacio intercelular.
Los desmosomas, también llamados macula adherens, son otro de los tipos de uniones que encontramos entre las células aracnoideas. En los epitelios, estas uniones permiten unir membranas de varias células adyacentes en un solo punto 28-30 . Estas uniones son habitualmente abundantes en zonas donde se producen tensiones mecánicas. En los desmosomas se identifica una placa densa adosada a la cara citosólica de cada una de las membranas plasmáticas que participan en la unión 28-30 .
Los desmosomas, a diferencia de las uniones estrechas, no adoptan la forma de cintas. Tienen dos placas de anclaje, de unos cientos de nanómetros de forma circular, con un espesor 10 a 15 nanómetros, formada al menos por 12 tipos de proteínas. Las dos membranas plasmáticas en esa zona tienen su espesor habitual y están separadas por 20 manómetros. Estas placas están formadas por glucoproteínas integrales, de la familia de las cadherinas. La placa contiene placoglobina y desmoplaquina I y II. Estas proteínas fijan la placa al citoesqueleto y a filamentos, denominados filamentos intermedios con 10 nanómetros de espesor, los que forman un lazo en horquilla por fuera de la placa, alejándose del desmosoma.
Durante años se ha mantenido que la duramadre era el principal componente en el control de la difusión a través del saco dural, si bien es una estructura totalmente permeable. Se podría suponer que el adelgazamiento que muestra la duramadre en la zona de los manguitos durales, podría facilitar la difusión de sustancias en esa zona. No obstante, probablemente el efecto barrera en esa zona es independiente del espesor de la duramadre y estaría aportado por un plano celular de transición 26 , con fuertes uniones especializadas de membrana, similares a las encontradas entre las células aracnoideas de la lámina aracnoidea. El importante papel que se ha atribuido a la duramadre en relación con la difusión, mantenido durante años, probablemente puede explicarse por su mayor grosor, ya que representa aproximadamente el 90% del espesor del saco dural, mientras que la lámina aracnoidea es muy delgada, transparente, frágil y se rompe fácilmente durante su manipulación 32 . Nuestros resultados permiten comprobar que no toda la lámina aracnoidea actúa como barrera. Las características ultraestructurales de la capa barrera ubicada dentro del espesor de la lámina aracnoides sugieren, por el contrario, que sólo este plano celular de la lámina aracnoidea es el factor que limita el paso de sustancias. El estrecho y tortuoso espacio que existe entre las células aracnoideas de apenas 20 a 30 nanómetros, interrumpido de forma frecuente por desmosomas y uniones estrechas, obligan a un paso transcelular y a un dificultoso tránsito intercelular de las sustancias. El gran número de vesículas de pinocitosis que se pueden observar en ambas superficies externa e interna de las células aracnoideas de la capa barrera, apoyan también esta interpretación. Bernards et al a partir de sus estudios de permeabilidad del saco dural 10-12 , postularon que las células aracnoideas podrían controlar hasta el 90% del paso de sustancias a través del mismo.
La permeabilidad meníngea espinal a sustancias lipofílicas puede determinarse según el coeficiente de distribución octanol-buffer 33-34 . La morfina tiene un valor octanol-buffer de 1, la lidocaína de 110, el alfentanilo de 129, la bupivacaína de 560, el fentanilo de 955, y el sufentanilo de 1.737. Si, por el contrario, se ordenan estos fármacos de acuerdo con su coeficiente de permeabilidad, de mayor a menor, se encuentra primero el alfentanilo (2,30 cm / minuto x 10 –3 ), seguido por la bupivacaína (1,60), lidocaína (1,45), fentanilo (0,92), sufentanilo (0,75), y por último, la morfina (0,62).
Curiosamente, las moléculas con un coeficiente intermedio de octanol:buffer son las que presentan mayor permeabilidad a través del saco dural. Las sustancias muy hidrofílicas, con un coeficiente de distribución octanol:buffer igual a 1, como la morfina, casi no tienen diferencias de permeabilidad con respecto a sustancias con un coeficiente de distribución octanol: buffer 1.737 como el sufentanilo.
Las características estructurales de la aracnoides y de la duramadre podrían justificar, por qué moléculas de solubilidad intermedia en lípidos son más permeables que otras moléculas que son muy liposolubles o muy hidrosolubles.
En un paso transcelular, las moléculas lipofílicas atravesarían con mayor facilidad las membranas plasmáticas con alto contenido en lípidos, pero retrasarían su paso al atravesar el medio acuoso intracelular. Con las moléculas hidrosolubles ocurría lo contrario. Probablemente, las moléculas con propiedades de solubilidad intermedia tengan mayor facilidad para atravesar sucesivos medios acuosos y lipídicos. Por otra parte, la polaridad de los componentes de la superficie externa de la membrana de las células aracnoideas, probablemente, interactúe con la densidad de cargas de moléculas, facilitando o dificultando su paso a través del estrecho espacio intercelular.
El paso de sustancias a través del saco dural se produciría por difusión simple, a favor del gradiente de concentración establecido entre ambos lados de la lámina aracnoidea. En estos casos, el movimiento de las moléculas respondería al movimiento browniano y al grado de partición dentro del medio donde termodinámicamente estas moléculas se encuentren más estables. Por esta razón, el patrón de partición jugará un rol determinante en la biodisponibilidad para que una molécula llegue a su sitio. Los múltiples vasos que hay en el espesor de la duramadre, compartimento subdural y piamadre, será otro factor a considerar, que disminuirá la llegada de fármacos a la médula, después de su administración epidural.
En el saco dural, la permeabilidad meníngea aumenta hasta alcanzar su valor máximo, próximo a un coeficiente de distribución octanol-buffer de 130, reproduciendo una relación bifásica como ocurre también en la piel y en la barrera hematoencefálica. Estos valores serían el reflejo de cómo las sustancias hidrofílicas y lipofílicas ajustan su paso a través de la aracnoides. En general, nuestros resultados coinciden con las observaciones de Vandenabeele 18 , aunque la morfología y tamaño celulares descritos muestran algunas diferencias. Nosotros encontramos que dentro de la lámina aracnoidea, las células aracnoideas de la capa barrera (tercera porción) no hacen contacto con las células neuroteliales (primera porción). Ambos planos celulares están separados por una membrana basal y un conjunto de fibras de colágeno, que daría cierta resistencia mecánica a esta membrana. Vandenabeele et al 18 , describieron una continuidad celular entre las células aracnoideas de la capa barrera y las células neuroteliales. La existencia de colágeno interpuesto entre ambas capas celulares, que a diferencia nosotros describimos, justificaría por qué el compartimento subdural se rompe siempre por el plano de las células neuroteliales, y no a través de las células aracnoideas de la capa barrera, algo más resistente, como ha sido descrito en trabajos previos 19,20 , en los cuales se estudió el origen del espacio subdural adquirido.
Entre las limitaciones del estudio debemos destacar el número de pacientes de los cuales se obtuvieron muestras. La posibilidad de conseguir este tipo de muestras es escasa dado el bajo número de pacientes con tumores de las raíces de la cola de caballo que se operan en nuestros hospitales. Además debe considerarse que no se forman fragmentos libres en todos los pacientes, sin que estos fragmentos estén afectados por la técnica quirúrgica, manteniendo una calidad óptima para su procesamiento. Cualquier alteración del tejido invalida la muestra para ser estudiada por microscopia electrónica de transmisión.
No obstante, a pesar del número de muestras, los hallazgos descritos se repitieron de forma homogénea en todas las imágenes analizadas.
En resumen, la lámina aracnoidea ocupa el 10% (35-40 μm) interno del total del saco dural (350 μm), y de esta lámina aracnoidea, sólo el 20% (5-8 μm) del espesor ocupado por un plano celular continuo de células aracnoideas asegura el efecto barrera de todo el conjunto, que justificaría la denominación de capa barrera dentro de la lámina aracnoidea. Las uniones especializadas de membrana entre estas células, obligarían a las moléculas a un paso transcelular, y en menor proporción, a un paso intercelular a través del estrecho desfiladero que limita el espacio intercelular.
1. Cousins MJ, Bromage PR. Bloqueo nerviosos epidural. En: Cousins MJ, Bridenbaugh PO. Bloqueos nerviosos. Barcelona: Doyma; 1991. p. 257-367.
2. Bromage PR. Anatomía. En: Bromage PR, editor. Anagesia epidural. Barcelona: Salvat; 1985. p.89-119.
3. Haberer JP. Anestesia peridural. En: Gauthier-Lafaye P. Anestesia locorregional. Barcelona: Masson; 1986. p. 165-205.
4. Churchill-Davidson HC. Analgesia espinal y epidural. En: Churchill-Davidson HC, editor. 3º ed. Barcelona:Salvat; 1983. p. 849-88.
5. Key A, Retzius G. Studien in der Anatomie des Nervensystems und des Bindegewebes. Estocolmo: Samson and Waller; 1875.
6. Bradford FK, Johnson PC. Passage of intact iron labeled erytrocytes from subarachnoid space to systemic circulation in dogs. J Neurosurg. 1962;19(4):332-6.
7. Alksne JF, Lovings ET. The role of arachnoid villus in the removal of red blood cells from arachnoid space. J Neurosurg. 1972;36(2):192-200.
8. Bernards CM, Hill HF. The spinal nerve root sleeve is not a preferred route for redistribution of drugs from epidural space to the spinal cord. Anesthesiology. 1991;75(5):827-32.
9. Bernards CM, Sorkin LS. Radicular artery blood flow does not redistribute fentanyl from epidural space to the spinal to the spinal cord. Anesthesiology. 1994;80(4):872-8.
10. Bernards CM, Hill HF. Physical and chemical properties of drug molecules governing their diffusion through the spinal meninges. Anesthesiology. 1992;77(4):750-6.
11. Bernards CM, Hill HF. Morphine and alfentanyl permeability through the spinal dura, arachnoid and pia mater of dog and monkeys. Anesthesiology. 1990;73(6):1214-9.
12. Bernards CM, Ulma G, Kopacz DJ. The meningeal permeability of rand S-bupivacaine are no different. Anesthesiology. 2000;93(3):896-7. 13. Reina MA, López A, Dittmann M, De Andrés JA. Análisis de la superficie externa e interna de la duramadre humana por microscopía electrónica de barrido. Rev Esp Anestesiol Reanim. 1996;43(4):130-4.
14. Reina MA, Dittmann M, López A, van Zundert A. New perspectives in the microscopic structure of human dura mater in the dorso lumbar region. Reg Anesth. 1997;22(2):161-6.
15. Reina MA, López A, Dittmann M, De Andrés JA. Análisis estructural del espesor de la duramadre humana por microscopía electrónica de barrido. Rev Esp Anestesiol Reanim. 1996;43(4):135-7.
16. Dittmann M, Reina MA, López A. Neue ergebnisse bei der darstellung der dura mater spinalis mittles rasterelektronenmikroskopie. Anaesthesist. 1998;47(5):409-13.
17. Reina MA, Pulido P, López A. El saco dural humano. Rev Arg Anestesiol. 2007;65(3):167-84.
18. Vandenabeele F, Creemers J, Lambrichts I. Ultrastructure of the human spinal arachnoid mater and dura mater. J Anat. 1996;189(2):417-30. 19. Reina MA, De León Casasola OA, López A, De Andrés JA, Mora M, Fernández A. The origin of the spinal subdural space. Ultrastructure finding. Anesth Analg. 2002;94(4):991-5.
20. Reina MA, López A, De Andrés JA, Villanueva MC, Cortés L. ¿Existe el espacio subdural?. Rev Esp Anestesiol Reanim. 1998;45(9):367- 76.
21. Reina MA, Villanueva MC, López A. Aracnoides trabecular, piamadre espinal humana y anestesia subaracnoidea. Rev Arg Anestesiol. 2008;66(2):111-33.
22. Reina MA, Machés F, López A, De Andrés JA. The ultrastructure of the spinal arachnoid in humans and its impact on spinal anesthesia, cauda equina syndrome and transient neurological síndrome. Tech Reg Anesth Pain Manage. 2008;12(3):153-60.
23. Reina MA, López A, De Andrés JA. Hipótesis sobre las bases anatómicas del síndrome de cauda equina e Irritación Radicular Transitoria post anestesia espinal. Rev Esp Anestesiol Reanim. 1999;46(3):99- 105.
24. Reina MA, De Leon Casasola O, Villanueva MC, López A, Maches F, De Andrés JA. Ultrastructural findings in human spinal pia mater in relation to subarachnoid anesthesia. Anesth Analg. 2004;98(5):1479- 85.
25. Reina MA, López A, De Andrés JA. Perforaciones naturales en la piamadre humana. Rev Esp Anestesiol Reanim. 1998;45(1):4-7.
26. Reina MA, Villanueva MC, Machés F, Carrera A, López A, De Andrés JA. Ultrastructure of human spinal root cuffs in lumbar spine. Anesth Analg. 2008;106(1):339-44.
27. Reina MA, Villanueva MC, López A, De Andrés JA. Grasa dentro de los manguitos durales de las raíces nerviosas de la columna lumbar humana. Rev Esp Anestesiol Reanim. 2007;54(5):169-72.
28. Junqueira LC, Carneiro J. Histología básica. 6º edición. Barcelona:Masson; 2006. p.67-91.
29. Kessel RG. Basic Medical histology. The biology of cells, tissues and organs. New York: Oxford Universtity; 1998. p 72-102.
30. Stevens A, Lowe J. Histología humana. Madrid: Elsevier; 2006. p. 39-46.
31. Genneser F. Histologia 3º ed. Buenos Aires: Editorial Médica Panamericana; 2000. p. 160-74.
32. Bernards CM. Sophistry in medicine: Lessons from the epidural space. Reg Anesth Pain Medicine. 2005;30(1):56-66.
33. Strichartz GR, Sánchez V, Arthur GR, Chafetz R, Martín D. Fundamental properties of local anesthetics: II. Measured octanol: buffer partition coefficients and pKa values of clinically used drugs. Anesth Analg. 1990;71(2):158-70.
34. Mather LE. Clinical pharmacokinetics of fentanyl and newer derivatives. Clin Pharmacokinet. 1983;8(5):422-46.
