Conectividad funcional y redes complejas en el estudio de la epilepsia focal.
Implicaciones fisiopatológicas y terapéuticas
Jesús Pastor, Rafael G. Sola, Lorena Vega-Zelaya, Óscar Garnés, Guillermo J. Ortega
Servicio de Neurofisiología Clínica (J. Pastor, L. Vega-Zelaya, O. Garnés); Servicio de Neurocirugía (R.G. Sola, G.J. Ortega); Instituto de Investigaciones Biomédicas (R.G. Sola, G.J. Ortega). Hospital Universitario de La Princesa. Madrid, España.Correspondencia:
Dr. Guillermo J. Ortega Rabbione. Fundación de Investigación Biomédica. Hospital Universitario de La Princesa. Diego de León, 62. E-28006 Madrid.
Introducción. El modelo tradicional de cirugía de la epilepsia focal consiste en la localización y resección de la zona epileptógena. Sin embargo, una minoría significativa de pacientes epilépticos sigue teniendo crisis tras la resección, lo que muestra el nivel de conocimiento incompleto que actualmente se tiene sobre esta materia.
Objetivo. Introducir algunos conceptos de conectividad funcional y redes complejas y su aplicación al estudio de registros neurofisiológicos de pacientes con epilepsia focal farmacorresistente, principalmente del lóbulo temporal.
Desarrollo. Con el objeto de entender con plenitud los nuevos desarrollos en el área de redes complejas y sus aplicaciones al estudio de la epilepsia, se revisarán los conceptos fundamentales de la metodología de redes complejas, sincronización y conectividad funcional. Asimismo, se revisarán algunos de los trabajos más recientes en los que se analizan registros neurofisiológicos y resonancias funcionales de pacientes con epilepsia focal bajo esta nueva perspectiva.
Conclusiones. Una visión más amplia en la perspectiva para el entendimiento de la epilepsia focal, como la tratada en este trabajo, permitirá también plantear nuevas alternativas al tratamiento quirúrgico de la epilepsia focal, tanto con el desarrollo de nuevos métodos no invasivos como con la mejora y optimización de algunos de los métodos invasivos ya existentes.
Palabras clave. Electroencefalografía. Epilepsia del lóbulo temporal. Redes límbicas. Sincronización. Teoría reticular.
Introducción
El tratamiento quirúrgico de la epilepsia farmacorresistente es una opción terapéutica válida para controlar (paliativa) o para eliminar (curativa) las crisis epilépticas en pacientes con epilepsia focal, especialmente en aquellos casos cuyo foco epileptógeno se sitúa en el lóbulo temporal (ELT).
En la epilepsia focal se asume que las descargas se originan en áreas específicas de la corteza cerebral –conocidas como zonas de comienzo ictal (ZCI)– desde donde la actividad eléctrica patológica se propaga a otras áreas de la corteza –como la zona sintomatogénica (ZS)– y a estructuras profundas. La relación entre estas regiones y la zona epileptógena (ZE), cuya definición operacional implica la desaparición de las crisis, no está bien establecida. La capacidad de propagación de una crisis comicial depende de la ZE [1,2], que en ocasiones coincide con la ZCI. En la actualidad, la resección de la ZE –habitualmente conocida como foco epiléptico– es el procedimiento más eficaz para disminuir o eliminar el número de crisis en la epilepsia farmacorresistente del lóbulo temporal. Sin embargo, la interacción real entre estas áreas sigue siendo objeto de debate [3]. De manera tradicional, se asocia una única área anatómica patológica como la responsable de la génesis de las crisis epilépticas asociada a alteraciones en el hipocampo y en la corteza entorrinal (atrofia, fundamentalmente) [4,5].
En los últimos años, se ha cuestionado el modelo focal –o de áreas– descrito en el párrafo anterior como paradigma explicativo del origen de la epilepsia focal [6], y se ha argumentado a favor de un modelo de redes en el que el énfasis ya no estaría en el foco o focos epilépticos, sino en las propiedades intrínsecas de la red cortical.
Aun cuando la cirugía resectiva de la epilepsia del lóbulo temporal es un tratamiento eficaz, es discutible afirmar si tal éxito terapéutico es la consecuencia de la resección del foco epileptógeno o, por el contrario, de una alteración en la topología de la red cortical en la que participaba la zona resecada. De hecho, en una minoría significativa de pacientes en los que se localiza la ZE durante los estudios preoperatorios, no se obtiene el éxito deseado tras la resección quirúrgica, argumento que va a favor del concepto de ‘patología de la red epiléptica’, si bien es cierto que hay otras posibles explicaciones de este fracaso terapéutico (resección insuficiente o defectuosa, diagnóstico incorrecto, complicaciones postoperatorias, existencia de diferentes umbrales epileptógenos, por citar sólo unas cuantas posibilidades).
El análisis de la relación espacio-tiempo entre la actividad ictal y la aparición de descargas epileptógenas interictales (DEI) ha dado lugar, por otro lado, a numerosos estudios que sugieren que la identificación y resección de aquellas regiones implicadas en la generación de DEI mejoran el resultado quirúrgico, pues permiten resecciones más ajustadas a la ZE [7]. Estos estudios describen la morfología de los episodios paroxísticos interictales bien como primarios o bien como actividad propagada, y tratan de caracterizar la distribución espaciotemporal en el registro electroencefalográfico (es decir, el origen de la actividad interictal y su patrón de propagación). Bajo esta perspectiva, Bourien et al propusieron un método en 2005 que determina automáticamente subestructuras implicadas en la generación de la actividad interictal sincronizada, y trataron de correlacionarlas con otras subestructuras implicadas en la génesis de la actividad ictal [8]. La importancia en la identificación de las descargas DEI simultáneas en distintas estructuras límbicas ya había sido observada por Spencer y Spencer en 1994 [9] y, desde entonces, se había prestado especial atención a la sincronización de la actividad paroxística interictal en el análisis de los registros neurofisiológicos provenientes de pacientes con ELT. Los dos enfoques antes descritos, la ELT como una alteración de la red límbica y la sincronización de actividad epileptogénica como una señal patológica, se han fusionado en años recientes y han dado origen a toda una nueva perspectiva desde la que se estudia actualmente la epilepsia. El desarrollo simultáneo de lo que en la actualidad se denomina teoría de redes complejas [10-13] ha potenciado la aplicación de esta nueva metodología en todo tipo de registros neurofisiológicos, de resonancia magnética funcional (RMf ) y magnetoencefalografía (MEG) provenientes de pacientes con epilepsia [1417] con el objeto de descubrir, por una parte, la estructura y funcionalidad de la red límbica –en el caso de la ELT– y, por la otra parte, el papel de la sincronización y desincronización en estos pacientes..
La metodología de redes complejas ha motivado que se vuelvan a analizar desde una perspectiva distinta los datos provenientes de pacientes con ELT
y también ha ayudado a la construcción de nuevos modelos numéricos y conceptuales sobre la base de los cuales es posible interpretar los hechos básicos de esta enfermedad desde una perspectiva diferente a la del modelo focal orientado a la identificación de áreas específicas, ZE, ZCI, entre otros.
En este trabajo se revisan brevemente los conceptos de sincronización y redes complejas y algunas de sus aplicaciones al estudio de las epilepsias parciales, fundamentalmente de la ELT farmacorresistente. Esta metodología se aplica por lo general y de modo retrospectivo sobre los diversos tipos de registros neurofisiológicos, electroencefalografía (EEG), electrocorticografía (ECoG), EEG semiinvasiva con electrodos de foramen oval (EFO) e invasiva con electrodos profundos (EP), y también en los estudios de RMf y MEG de pacientes diagnosticados con epilepsia parcial. Mostraremos en la parte final de esta revisión algunas propuestas alternativas para el abordaje terapéutico de la epilepsia parcial basadas en esta nueva visión.
¿Sincronización de actividad epileptogénica o de actividad basal?
Antes de describir los conceptos básicos de sincronización y redes complejas es conveniente resaltar la diferencia fundamental existente en el estudio de registros electrofisiológicos interictales, ya sea desde un punto de vista tradicional o desde un punto de vista de redes complejas. Clásicamente, el énfasis ha estado puesto sobre la actividad anormal epileptogénica [18,19] y el concepto de sincronización ha sido aplicado a la aparición simultánea de este tipo de actividad en registros de distintas estructuras límbicas como indicación de interacción patológica [8,9] entre ellas. Como veremos en el apartado siguiente, la forma usual de cuantificar la existencia, o no, de sincronización entre áreas corticales registradas por distintos métodos electrofisiológicos es a través de la señal basal (o de fluctuaciones espontáneas en el caso de la RMf) sin tener en cuenta la presencia de actividad epileptogénica. No obstante, es posible que una alta actividad paroxística en dichos registros altere el valor de sincronización de las señales basales. Debido a este hecho, en algunos casos [20] es necesario eliminar de los registros dicha actividad interictal, fundamentalmente la debida a las DEI. Hace poco, hemos demostrado [21, 22] que en la mayoría de los casos no es necesario eliminar la actividad paroxística de los registros neurofisiológicos a fin de obtener valores de sincronización de la actividad basal, salvo en el caso de registros provenientes de EP, que contienen una gran cantidad de puntas interictales y la sincronización de ellas puede afectar sensiblemente el valor final de sincronización. En todo caso, siempre deberán evaluarse los valores de sincronización promedios obtenidos y los tipos de electrodos empleados (EEG, ECoG, EFO, entre otros) con el objeto de estimar la cantidad de contaminación de la sincronización de la actividad epileptogénica sobre la actividad basal [21].
Sincronización y conectividad funcional
El concepto de sincronización hace referencia, en términos muy generales, a la coincidencia temporal de dos o más eventos. Desde la primera descripción del fenómeno de sincronización hecha por Huygens en 1673 hasta nuestros días, el concepto de sincronización ha evolucionado y se ha diversificado de muchas formas [23-26]. En lo que se refiere a este trabajo, usaremos la noción de sincronización comúnmente empleada y que puede expresarse diciendo que dos sistemas se encuentran sincronizados si sus respectivos ritmos (oscilaciones) se ajustan debido a la interacción existente entre ellos. En ese sentido, decimos, por ejemplo, que dos áreas corticales distantes se encuentran sincronizadas si la actividad rítmica de cada una de ellas, registradas por ejemplo con electrodos de ECoG, es similar debido a una interacción (directa o indirecta) entre ellas. A la hora de cuantificar la similitud entre oscilaciones se ha propuesto una gran cantidad de medidas. Una de las más empleadas actualmente es el coeficiente de correlación de Pearson, medida de asociación lineal entre dos series temporales de N elementos cada una, x i y x j , para un tiempo k :
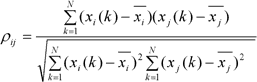
donde ρ ij es la correlación lineal entre las series de tiempo del nodo i y la serie de tiempo del nodo j . Actualmente, también se emplea mucho la sincronización de fase; esta medida es una importante generalización de la función de coherencia, un coeficiente de correlación en el dominio de frecuencias, empleado para tratar con osciladores no lineales y sistemas estocásticos.
En el área de las neurociencias en particular, el concepto de sincronización fue reformulado por Friston en 1993 [27] de tal modo que si dos o más áreas corticales se encuentran sincronizadas se dice que entre las mismas existe una conectividad funcional (CF) que las relaciona [28]. Este tipo de conectividad puede o no estar relacionada [29,30] con la conectividad neuroanatómica o estructural (CE), que muchas veces es difícil de identificar. Un ejemplo de conectividad neuroanatómica a escala macroscópica es la de los tractos y fascículos de la sustancia blanca identificados en las tractografías. Por último, cuando es posible evaluar la causalidad existente entre la actividad de dos zonas corticales, se define la conectividad efectiva para determinar qué área manda y que área obedece. Puede consultarse el reciente trabajo de van Diessen et al [31] sobre las relaciones entre CF y CE en la epilepsia. Puesto que la estimación de los valores de sincronización invariablemente arroja un resultado numérico, entre 0 (falta de sincronización) y 1 (sincronización perfecta), muchas veces es necesario establecer umbrales para determinar cuándo ese valor es representativo de una CF. Por ejemplo si los valores de sincronización calculados son menores a 0,2, entonces podemos decir que no existe CF entre las zonas en cuestión. Realizando lo mismo para todas las estimaciones de correlación entre los nodos de la red obtendremos su patrón de conectividad, o la topología funcional. Como veremos más adelante, esta información resulta fundamental para caracterizar la red. En la figura 1, por ejemplo, podemos observar la estimación de la CF entre tres zonas corticales simuladas X , Y y Z .
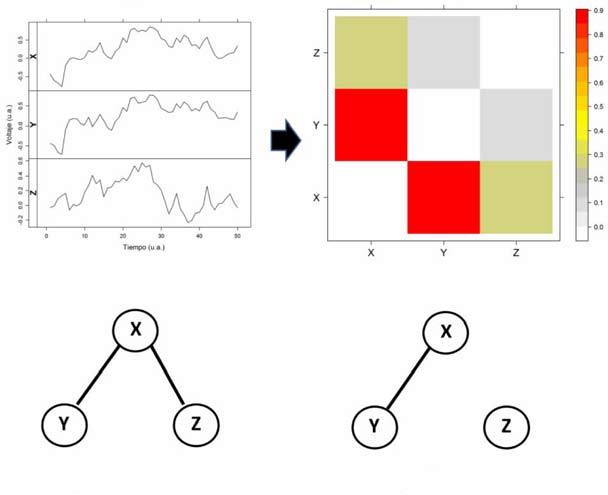
La actividad bioeléctrica puede estar representada por un EEG o un ECoG, entre otros, que se representa en la figura 1a para las tres áreas corticales. Usando la ecuación 1 para todos los pares, podemos obtener la matriz de correlación representada en la figura 1b. En ese caso, como en los siguientes, se ha usado el valor absoluto de la correlación calculada, de tal modo que la variación es entre 0 (sin correlación) y 1 (correlación o anticorrelación perfecta). Claramente puede observarse, tanto en la parte A de los registros como en la matriz de correlación B, la alta correlación existente entre los registros X e Y , como así también la menor correlación entre X y Z e Y y Z .
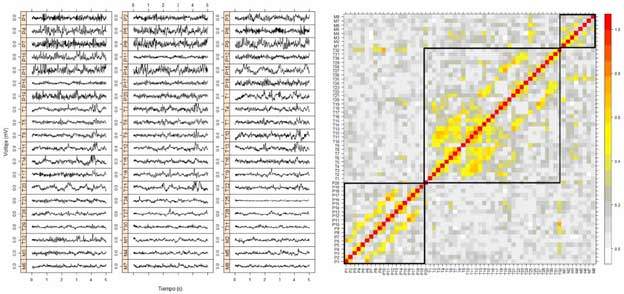
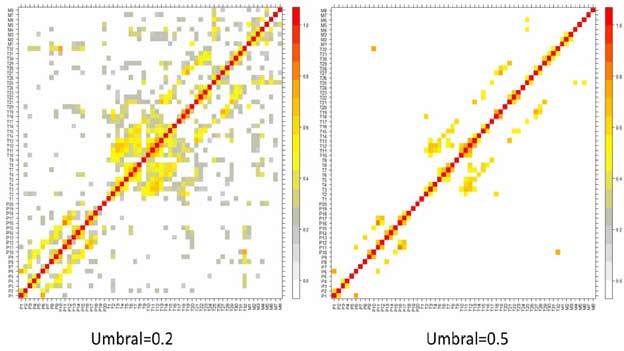
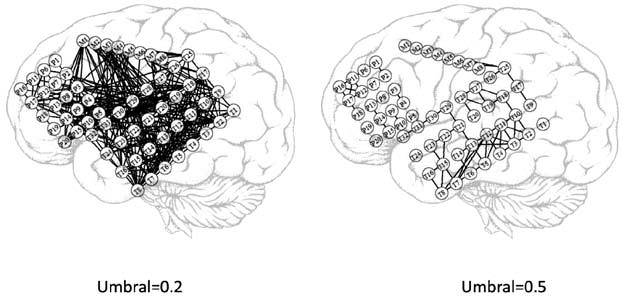
A la hora de determinar la conectividad, de acuerdo con lo explicado más arriba, es fundamental la determinación del umbral para descartar aquellas interacciones que pueden considerarse débiles. Por ejemplo, en la figura 1c puede verse la determinación de la CF a partir de la matriz de correlación usando un umbral de 0,3 para las correlaciones significativas, esto es, aquellas correlaciones menores a 0,3 no se consideran, por lo que habrá CF entre las áreas X e Y y X y Z , como se muestra en la figura 1c. Alternativamente, cuando el umbral se aumenta a 0,6, sólo queda presente la conectividad entre X e Y , como se muestra en la figura 1c. De este modo, el área Z quedaría desconectada de las otras áreas. Un ejemplo real puede observarse en la figura 2, obtenido por medio de ECoG crónicas con electrodos subdurales en el caso de un paciente con epilepsia extratemporal. En este caso, se han implantado 60 electrodos distribuidos en tres mantas que cubren partes de las zonas parietales (P1-P20), temporal lateral (T1-T32) y temporal mesial (M1-M8). En la figura 2, pueden observarse los registros de los 60 canales (parte A) durante 5 segundos. En la figura 2b, puede observarse la matriz de correlación, calculada con el valor absoluto de la ecuación 1, donde se han recuadrado en negro las zonas de interacciones intraáreas. El patrón de correlaciones puede distinguirse con claridad. Esto es más evidente aún al usar umbrales. Por ejemplo, usando un umbral de 0,2, puede observarse en la figura 3a que la CF aparece principalmente como interacciones interzonas, con algunas excepciones. Usando un umbral más alto, por ejemplo igual a 0,5, puede observarse en la figura 3b que sólo quedan las interacciones más importantes. En la figura 4, podemos observar una representación aproximada de la CF de estos electrodos subdurales con su localización espacial sobre la corteza cerebral. La tira de electrodos mesiales (M1-M8), que está situada en la parte interna del lóbulo temporal, se ha representado en una zona que no le corresponde solamente para mejorar su visualización. En la figura 4a, se representa la matriz de correlación con un umbral de 0,2 (Fig. 3a) y, en la figura 4b, la correspondiente a un umbral de 0,5 (Fig. 3b). Se puede observar en esta última figura que las interacciones son predominantemente intrazonas, salvo en el caso de la interacción lateralmesial, a través del electrodo T25 y la interacción parietal-temporal a través de la CF T32-P10, esta última seguramente debido a la proximidad espacial entre ambos electrodos.
El procedimiento antes descrito, no obstante, tiene implícita la arbitrariedad en la elección del umbral, lo que requiere la exploración para distintos valores de umbrales [29]. Un procedimiento más natural empleado alternativamente consiste en el uso de árboles jerárquicos que permite estudiar las jerarquías en los valores de sincronización y los patrones de ordenamiento [32,33].
La determinación y cuantificación de la CF entre varias áreas corticales es el ladrillo fundamental en el que se basan las nuevas aproximaciones al estudio de los registros neurofisiológicos, ya que proporcionan uno de los dos elementos con los que están construidas las redes, esto es, las conexiones entre los nodos de las mismas.
Conectividad funcional y redes complejas
Una red compleja se define habitualmente en el contexto de la teoría de grafos, una rama de las matemáticas dirigida al estudio de un conjunto de nodos conectados entre sí. El objetivo al analizar las redes complejas es la descripción de las principales características de la red, la dinámica de sus nodos y sus enlaces o conexiones, tarea que no puede hacerse a simple vista. Hay muchos criterios para caracterizar una red compleja y los elementos que la conforman, nodos y enlaces. Una manera de caracterizar una red es por medio de la identificación de las comunidades o clusters de elementos contenidos en ella, de tal modo que cada comunidad individual dentro de una red reúne aquellos nodos que están más estrechamente ligados, o sincronizados, entre sí.
La identificación de aquellos nodos de la red que desempeñan un papel sobresaliente en la misma también se usa para caracterizar la red. De acuerdo con ello suelen definirse medidas de centralidad para los nodos de una red que permiten resaltar ciertas características, como por ejemplo en lo referente a la transmisión de información en toda la red. Dentro de las medidas de centralidad, resultan de especial interés, en lo referente a las redes epileptogénicas, los siguientes conceptos:
– Grado o conectividad de un nodo. Es el número de enlaces en los que participa un nodo. Para el caso de una red funcional, puesto que los enlaces están definidos por la sincronización, el grado da una idea de sincronización del nodo en cuestión con otros nodos de toda la red.
– Sincronización local (SL). Cuantifica el grado de sincronización entre un nodo y los inmediatamente más próximos. Para un nodo i , se define como el sumatorio de una medida de sincronización cualquiera (es decir, el valor absoluto del coeficiente Pearson) normalizado en función del número de nodos colindantes n ij :
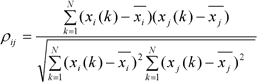
– Intermediación ( betweenness ) de un nodo i . Hace referencia a la frecuencia con la que ese nodo sirve de camino más corto entre otros dos nodos, y se define como el cociente entre el total de conexiones más cortas entre dos nodos cualesquiera de toda la red que pasan por el nodo i y el total de conexiones más cortas de la red que conectan ambos nodos. Dicho de otro modo, cuantifica la importancia de un nodo determinado dentro de la globalidad de la red. Una zona con nodos de alta intermediación se ha asociado con la zona epileptógena [14].
– Coeficiente de agrupamiento (CA) ( clustering coefficient ) o transitividad. Es la propiedad de un nodo de conectarse en triángulos con otros nodos. Esto es, si un nodo i se conecta con otro nodo j , que a su vez se conecta con otro nodo k , si el nodo i tiene un alto CA entonces es muy probable que el nodo k también esté conectado al nodo j . Se define, por lo tanto, como el cociente entre el número de conexiones reales y el número total de conexiones posibles entre un nodo y sus nodos colindantes.
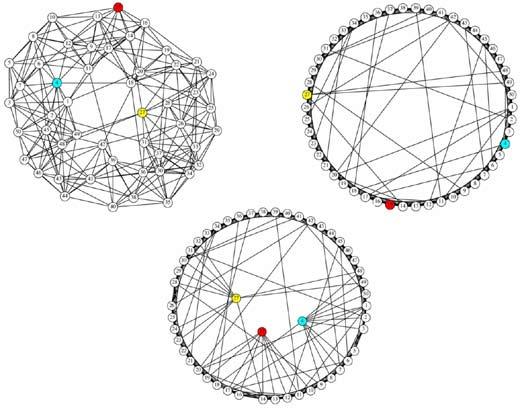
Por último, una medida importante, fundamentalmente en el análisis de las redes corticales, es la que se conoce como longitud del camino más corto ( shortest path length ) y que básicamente mide el camino más corto, en términos de trayectos entre nodos, que existe entre dos nodos cualesquiera de la red. A partir de esta medida suele definirse la longitud del camino más corto promedio como el promedio de la longitud del camino más corto para todos los pares de nodos de la red. Es evidente que, cuanto más pequeña sea la longitud del camino más corto promedio en una red, más rápida y, posiblemente, más eficiente sea la comunicación en la misma. Si a lo anterior se suma que una red posee nodos con alto CA, estaremos en presencia de lo que se conoce como la propiedad de mundo-pequeño ( small-world ), que permite transmitir información rápidamente entre nodos muy alejados debido a la presencia de caminos más cortos y de nodos con alto CA. En la figura 5, podemos observar una red arbitraria donde se han determinado algunas medidas de centralidad. En particular, se ha calculado para cada nodo el valor de CA, el grado y la intermediación. El máximo de cada una de estas medidas de centralidad se ha coloreado. Así, el nodo con máximo grado se ha pintado de color azul, el de mayor valor de intermediación, de color amarillo y el mayor valor de CA, de color rojo. Puede verse en este caso la importancia de los nodos con alto CA, ya que si bien el nodo rojo posee menos enlaces que el azul, no obstante está conectado con nodos que poseen enlaces a larga distancia y por lo tanto cualquier información que pase por él será rápidamente distribuida en toda la red. Veremos más adelante la importancia de estos nodos en lo que respecta a la distribución de información epileptogénica.
ELT mesial: ¿mayor o menor conectividad funcional?
Puesto que tradicionalmente se ha asociado la epilepsia a la sincronización [24,25], muchos de los estudios bajo la óptica de redes se han centrado en cuantificar la CF en pacientes con ELT. Los resultados no sólo no son concluyentes en esta cuestión, sino que la aparente contradicción entre ellos proviene del hecho de no ser comparables unos con otros. Por otro lado, algunos de ellos están realizados sobre registros neurofisiológicos y otros sobre RMf, que, si bien da una información indirecta sobre la actividad bioeléctrica, permite comparar contra auténticos casos controles. Respecto al análisis de registros electrofisiológicos interictales, Bettus et al [20], analizando registros de EP, han encontrado que existe una mayor CF en la zona mesial ipsilateral, en pacientes con ELT mesial, con respecto a registros de pacientes controles con epilepsia extramesial (lateral, frontal y occipital). Por otro lado, usando EFO, Ortega et al [32] han encontrado que la zona mesial, fundamentalmente la corteza entorrinal, presenta una menor CF que la contralateral en aquellos pacientes con ELT mesial. Por lo tanto, ambos estudios no muestran resultados contradictorios, ya que las comparaciones son distintas. Resultados con análisis de RMf de Liao et al [34] muestran que existe una mayor CF en la zona mesial respecto a un grupo control de personas sanas. Al contrario, y también usando RF, Pereira et al [35] han encontrado que existe una menor CF en la zona hipocampal en pacientes con ELT mesial, con esclerosis de hipocampo. Resultados similares han sido descritos por Bettus et al [36].
ELT y heterogeneidad en la conectividad funcional
En el apartado anterior, hemos mostrado algunos resultados relacionados con la sincronización global en la zona mesial de pacientes con ELT. Desde el punto de vista local, no obstante, ha habido varios resultados semejantes entre sí [14,37-41] en lo relativo a la distribución espacial de la CF en estructuras límbicas en pacientes con ELT, y quizás sea éste uno de los resultados más robustos que existen actualmente en lo referente al análisis de registros neurofisiológicos en pacientes con epilepsia [42]. En estos trabajos, se ha demostrado que la CF no está distribuida de modo uniforme en la corteza cerebral de estos pacientes. En el caso de pacientes con ELT, esta inhomogeneidad aparece tanto en la corteza lateral del lóbulo temporal como en la zona mesial. No obstante, la relación de estas zonas de alta CF con la zona de inicio ictal es una cuestión no aclarada. Mientras en algunos casos se ha mostrado que hay una relación entre las mismas [41], en otros no ha podido establecerse una relación directa. Una cuestión importante en lo relativo a estas zonas es su estabilidad temporal. En un trabajo reciente hemos mostrado [38] que estas zonas de alta CF muestran a su vez una estabilidad temporal muy alta, por lo que su papel en la dinámica cortical es doble al ser zonas de alta CF muy estables en el tiempo, lo que haría a la red cortical más propensa a entrar en un estado de sincronización global, esto es, propensa a las crisis ( seizurability ). Por otro lado, también hemos mostrado [37] que el grado de localización de estas zonas se relaciona con los resultados de las cirugías resectivas. Cuanto más concentrada espacialmente está la CF y dicha zona es resecada durante la cirugía, mejor pronóstico tiene el paciente.
Redes y actividad ictal
La descripción de la dinámica ictal en pacientes con epilepsia es una cuestión fundamental que también ha sido abordada por la nueva metodología. Se ha descrito un aumento simultáneo del CA, de la longitud del camino más corto y de la densidad de enlaces, esta última una medida de la sincronización global, durante las crisis con respecto a la actividad interictal [43-46]. Los resultados anteriores muestran, por lo tanto, que existe una reestructuración de la red funcional hacia una estructura más organizada al hacer la transición interictal-ictal. De hecho, se ha mostrado que durante las crisis mismas hay una estructura interna en lo que respecta a las variaciones de estas medidas, lo que podría dar claves fundamentales sobre los mecanismos de inicio y, quizás más importante aún, de la finalización de las crisis. En el caso de las crisis focales, Kramer et al [44] han mostrado que al inicio de las crisis existe una única red muy regular, que, a medida que la crisis avanza, tiende a fragmentarse en varias subredes menores, hasta llegar a la finalización de la crisis. El llamado efecto de mundo-pequeño también ha sido estudiado durante las crisis, y se ha encontrado que hay un aumento de esta propiedad [43,47] durante las crisis, lo que produciría una mayor facilidad de comunicación entre todos los nodos de la red y, por lo tanto, facilitaría la propagación de las crisis. No obstante, estos resultados han sido discutidos en algunas publicaciones [48-50].
Implicaciones clínicas del enfoque de redes
El paradigma de una epilepsia reticular o distribuida frente al clásico de epilepsia focal tiene implicaciones tanto diagnósticas como terapéuticas que van más allá del estudio fisiopatológico de la epilepsia. La evaluación prequirúrgica de la epilepsia farmacorresistente, según Spencer, debería estar orientada a la caracterización de la red epiléptica, en lugar de estudiar y localizar, como se hace actualmente, zonas particulares de la corteza cerebral, esto es, la ZE y la ZCI [6]. El conocimiento y práctica adquiridos durante décadas bajo la aproximación clásica –orientada a la determinación de las ZE y ZCI– tiene que servirnos en el presente para realizar una transición suave hacia la nueva visión de la epilepsia como una enfermedad de la red cortical afectada, fundamentalmente en lo que respecta al tratamiento quirúrgico. Un primer paso en este sentido ha sido llevado a cabo por nuestro grupo al diferenciar dos tipos de comportamiento bioeléctrico en la ECoG intraoperatoria, focal o reticular, y que ha permitido modificar el procedimiento quirúrgico, minimizando la región resecada y, por lo tanto, ofreciendo un tratamiento individualizado para cada paciente en función del sustrato fisiopatológico de su epilepsia [51,52].
Conectividad funcional y redes complejas en el estudio de la epilepsia focal
Functional connectivity and complex networks in focal epilepsy. Pathophysiology and therapeutic implications
Introduction. The traditional surgical approach to treat drug-resistant focal epileptic patients is in the resection or disconnection of the epileptic focus. However, a significant minority of patients continue to experience seizures after surgery, which shows the incomplete level of knowledge that currently we have of this pathology.
Aim. This paper introduces some concepts of functional connectivity and complex networks methodology with its application to the study of neurophysiological recordings from patients suffering from drug-resistant focal epilepsy.
Development. In order to fully understand the new developments in the area of complex networks and its applications to the study of epilepsy, we will here review fundamental concepts in complex networks methodology, synchronization and functional connectivity. Some of the most recent published works dealing with focal epilepsy viewed under this new perspective will be revised and commented.
Conclusions. We think that a wider perspective in the study of epilepsy, such as the one reviewed in this work, will allow epileptologists to consider surgical alternatives in the usual treatment of focal epilepsy at those currently performed in most medical centers around the world. Combining the traditional knowledge with new insights provided by network theory will certainly fill many of the gaps we have today in the fragmented understanding of epilepsy.
Key words. Complex network. Electroencephalography. Functional connectivity. Synchronization. Temporal lobe epilepsy.

muy buen tema